19 Proto- and Early Hominins
Tori M Saneda

Our Earliest Ancestors
In determining what fossil features denote a specimen is a hominin, many different characteristics are examined, including those related to bipedalism, about which you previously learned, and non-honing chewing. Apes have a canine-premolar honing complex, which means that there is a diastema between the lower canine lower third premolar where the upper canine fits when the jaws close (Larsen 2014). Other characteristics such brain and body size and what, if any, cultural behaviors are associated with the fossil remains are also scrutinized.
The various features associated with hominins developed at different rates, a situation referred to as mosaic evolution. In determining which species a fossil specimen belongs to, paleoanthropologists utilize comparative anatomy. There is much discussion among researchers as to the exact number of species. Some tend to see broad ranges of variation, which results in fewer species. Researchers with this approach are sometimes referred to as “lumpers” because they lump fossil specimens in relatively few species. Other researchers see more narrow ranges of variation and are referred to as “splitters” because they see more variation of species in the fossil record. For the purposes of this course, we are going to use the approach presented in Jurmain et al. (2013), which is more of a lumper approach.
While the hominins will be presented more or less in chronological order, do not mistake chronological order for linear evolutionary relationships; some hominins that are presented are not in the direct line to modern humans. It is also important to keep in mind that new discoveries are made each year that refine what we know about human evolution. What this means is that what you learn this quarter could change somewhat next year as new data is made available. While this can make studying human evolution frustrating at times, the new discoveries help us gain a better understanding of just how our early ancestors evolved.
It is highly recommended that you begin your exploration of human evolution by watching the documentary, Becoming Human.
Proto-hominins
Sahelanthropus tchadensis
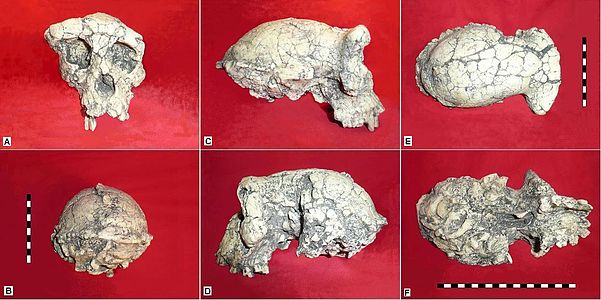
Discovered by the Mission Paléoanthropologique Franco-Tchadienne (MPFT) led by Michel Brunet at Toros-Menalla, Chad, during the 2001-2002 field season, Sahelanthropus tchadensis is the oldest hominin discovered to date. Cosmogenic nuclide dating (https://en.wikipedia.org/wiki/Surface_exposure_dating ) places Sahelanthropus between 7.2 to 6.8 mya (million years ago) (Lebatard et al. 2008). The binomial recognizes the area where the fossils were found, Sahel. The type specimen, also referred to as a holotype, includes “…a nearly complete cranium and…parts of two mandibles and some isolated teeth” (Bailey 2006: 2044).
The skull is a combination of ape-like and human-like features. Ape-like features include brain size, heavy brow ridge. Its human-like features include forward position of the foramen magnum (visit “The Earliest Hominins: Sahelanthropus, Orrorin, and Ardipithecus” (scroll down) to view image comparing placement of foramen magnum in chimps, humans, and Sahelanthropus), reduced prognathism, smaller canine teeth, and intermediate thickness of the premolar and molar enamel. Due to the pronounced brow ridge Brunet’s team suggests that the Sahelanthropus specimen is male.
There is debate among researchers as to whether Sahelanthropus is a hominin or an ape. Some suggest that the specimen belongs to that of a female ape because it is likely to find canines worn at the tips in female apes. Critics claim that without postcranial evidence it is premature to claim that Sahelanthropus is bipedal (Su 2013).
Paleoecological data suggest that this Sahelanthropus specimen died near a lakeshore and the area was extensively forested.
Orrorin tugenensis
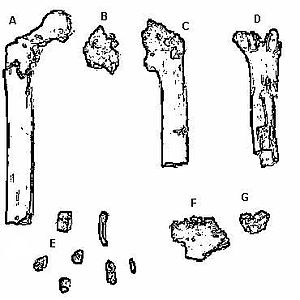
Found in 2000 by a team led by Martin Pickford and Brigitte Senut, Orrorin tugenensis is represented by a collection of fossils that include a minimum of five individuals. Fragments of numerous fossilized body parts were found: jaw, femur, humerus, and finger bones. Many teeth were found, including molars and canines. The binomial reflects where the fossils were found, the Tugen Hills region of Kenya. Orrorin means “original man” in the Tugen dialect. Using multiple dating techniques, including paleomagnetism and biochronology, the Tugen Hills fossils are dated to about 6.0 mya.
Like Sahelanthropus, Orrorin has a mix of ape-like and human-like traits. Its teeth are more ape-like so the determination of bipedality is central to attributing the fossils to that of a hominin. The founding research team claims that features of the femur indicate that Orrorin was a biped and in the direct line to modern humans. More recent studies suggest that Orrorin was in fact a bipedal hominin, but not in the direct line to Homo (Richmond and Jungers 2008).
The paleoecological evidence associated with Orrorin fossils suggests that the area was a woodland populated with lakes and streams.
Ardipithecus

Two different species of Ardipithecus have been identified: Ar. kaddaba and Ar. ramidus. The two species are differentiated primarily on the C/P3 complex (canine-premolar features), specifically, that Ar. kaddaba’s canines are more primitive than those of Ar. ramidus.
Ar. kaddaba is the older of the two species. It is dated to 5.8-5.2 mya. Tim White, Gen Suwa, and Yohannes Haile Selassie discovered Ar. kaddaba fossils in the Middle Awash region of Ethiopia. The fossil collection is comprised of fragments of the mandible and teeth, forearm, clavicle, and fourth toe. The shape of the upper canine is more human-like than ape-like in that they are smaller with a more circular crown. Overall, the postcranial fragments are more ape-like; however, the toe bone mentioned previously is similar to the toe bones of Australopithecus afarensis, a species that had all the hallmarks of habitual bipedalism. Other researchers claim that a single fourth toe bone does not provide enough evidence to confirm that Ar. kaddaba was a bipedal hominin.
Paleoecological data suggest that Ar. kaddaba lived in a “closed, densely-wooded habitat close to permanent water sources (e.g., lakes and/or rivers) with swampy conditions and floodplain grasslands” (Becoming Human c2008).
Tim White is also credited with the discovery of Ar. ramidus in Aramis, Ethiopia. Dated to about 4.4 mya, Ar. ramidus “represents the oldest species that possesses features unequivocally linked to the hominin lineage. Thus, Ar. ramidus is the best evidence discovered thus far for the root of the hominin family tree” (Becoming Human c2008). Paleoanthropologists know more about Ar. ramidus than the previously mentioned species as 110 specimens have been unearthed at Aramis.
Postcranially, Ar. ramidus is still ape-like; however there are some features that suggest the species could walk bipedally, but it most likely spent time in the trees much like modern chimpanzees. In other words, its mosaic characteristics suggest facultative bipedalism. Features that lead researchers to claim Ar. ramidus could walk bipedally include the position of the foramen magnum and features of the spine and ilium.
Early hominins
Australopithecus anamensis

Australopithecus anamensis, meaning “southern ape of the lake,” was found by Maeve Leakey and Alan Walker in 1994 in Kanapoi, Kenya. Additional Au. anamensis fossils were recovered at Allia Bay, Kenya and several sites in Ethiopia: Asa Issie, Woranso-Mille, and Fejej. Fossil specimens include mandibles, maxilla, teeth, a partial humerus, tibia, manual phalanx, capitate, partial femur, several vertebrae, and a partial temporal bone (Ward et al 2013). Using radioisotopic dating methods on volcanic deposits, Au. anamensis dates to 4.1-3.9mya. The fossil evidence suggests that Au. anamensis is ancestral to Au. afarensis.
Au. anamensis exhibits a mosaic of ape-like and Homo-like features. The size and shape of the teeth, especially the lower premolar and upper canine, the u-shaped dental arcade, and the prognathism are some of the ape-like features, while the thickness of the enamel and the breadth of the molars are more like early Homo. The single wrist bone is also like Homo. Lower limb bones suggest Au. anamensis was fully bipedal.
Paleoecological data suggest that Au. anamensis lived in forested areas near streams.
Australopithecus afarensis

Discovered by Donald Johanson in 1974 at Hadar, Ethiopia, Australopithecus afarensis is arguably the most well know fossil hominin species. It is dated from 3.7-3.0mya (Scarre 2014). Johanson’s discovery in 1974 was, at the time, the most complete hominin fossil. Over 40% of the skeleton was recovered, which allowed the team to fully reconstruct the skeleton. This fossil specimen, named Lucy, coupled with footprints found at Laetoli, Tanzania, in 1978 by Mary Leakey, confirmed that Au. afarensis was fully bipedal, albeit not exactly like modern humans. The footprints at Laetoli indicate that Au. afarensis had a short stride and a strolling gait. Since the 1970s, hundreds of specimens of Au. afarensis have been found (60 individuals at least from Hadar alone!) in Ethiopia, Kenya, and Tanzania, allowing paleoanthropologists to make “definitive [emphasis original] statements about the locomotor pattern and stature of” (Jurmain 2013: 211) this early hominin.
Au. afarensis has several primitive, or ape-like, features, including a relatively small brain in comparison to Homo, a u-shaped dental arcade, a flat nose, a flattened forehead (referred to as platycephaly) and prognathic face. Its canines, while larger than Homo, are smaller than earlier hominins. While its brain was larger than earlier hominins, it is still small in comparison to Homo. There is evidence for sexual dimorphism; Au. afarensis males were no taller than 5 feet and females about 3-4 feet, similar in proportion to modern African apes. Au. afarensis has a suite of derived traits associated with bipedalism; i.e., bowl-shaped pelvis, s-curve of the vertebral column, and knee anatomy. However, the curvature of the fingers and toes and the proportion of the arms to legs suggest to some researchers that Au. afarensis spent some time in the trees.
In 2006, Zeresena Alemsegad announced the discovery of a 3.3 million year old Au. afarensis child not 4km from where Lucy was found in 1974. It is not only the oldest juvenile fossil ever found, it is also the most complete hominin fossil found to date. Selam, as the fossil was nicknamed (or Lucy’s baby or Dikika’s baby), confirmed earlier suggestions that Au. afarensis was bipedal, yet spent time in the trees. The shoulder structure with its upward pointing shoulder joints and the bony ridge running along the shoulder blades are like that of apes, which would have facilitated arboreal movement even if they were not as fully capable as chimpanzees at moving in trees.
Paleoecological data indicates that Au. afarensis lived in both grassland (savanna) and woodland environments.
Australopithecus africanus
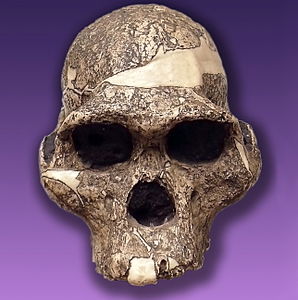
Raymond Dart identified Australopithecus africanus, “African southern ape,” in 1924, the first australopith to be recognized. This specimen was a juvenile australopith, nicknamed the Taung baby. Based on the position of foramen magnum, Dart claimed it was a bipedal hominin, but his contemporaries refuted his claim as they thought the brain was too small. The paleoanthropological community did not recognize the Taung baby as a hominin until the 1950s at which time other fossil specimens attributed to Au. africanus had been found. Au. africanus has only been found in southern Africa at such sites as Makapansgat, Sterkfontein, and Taung.
Using primarily biochronological methods since South Africa does not have geologic deposits suitable for other types of dating techniques, it has been dated to 3-2mya (Scarre 2014). In comparison to Au. afarensis, Au. africanus has smaller incisors and larger molars; the canines no longer have the pointed, triangular appearance seen in apes and Au. afarensis; however, Au. africanus still exhibits some prognathism in the face and has a small brain like that of Au. afarensis. Postcranially, Au. africanus retains some primitive features like those seen in Au. afarensis, e.g., shorter arms than legs and hand/feet shape. Its size is also similar to Au. afarensis. Another derived feature are the nasal pillars. Nasal pillars are a “buttressing of bone on either side of the nasal opening of the skill on the maxilla,” which may be an adaptation for eating hard foods (Becoming Human c.2008). Other derived traits include a longer, flatter occipital bone and a taller frontal bone, two traits seen in the genus Homo. Because of the traits mentioned above, there is general consensus that Au. africanus is a direct descendent of Au. afarensis. Some paleoanthropologists suggest that Au. africanus is the progenitor of Paranthropus robustus (see below). Its relationship to Homo is less clear.
Australopithecus garhi

Found in Bouri, Ethiopia in 1997 by Behane Asfaw and Tim White, Australopithecus garhi is a gracile australopith. “Gracile” refers to the thickness of the bone; in this case it is less robust than its Paranthropus contemporaries. Using radioisotopic and biochronometric dating methods, Au. garhi dates to 2.5mya. Few fossil specimens have been found and those that have are relatively fragmentary. One cranium and other skull fragments were found and serve as the basis of the species identification. The size and shape of its molar teeth suggest to some researchers that Au. garhi is related to Paranthropus aethiopicus (see below), but its other features, e.g., braincase, face, and other teeth, are more like genus Homo. In light of this, some researchers contend it is ancestral to Homo.
Australopithecus sediba

A relative newcomer to the stage, Australopithecus sediba was found in 2008 at Malapa Cave, South Africa by Matthew Berger, the 9-year-old-son of the project’s lead paleoanthropologist, Lee Berger. Dating to 1.95-1.75 mya using paleomagnetism, uranium-lead dating, and biochronology, Au. sediba has a mosaic of characteristics that suggest it may be transitional from the australopiths to genus Homo, hence its name “wellspring or fountain” in the local Lesotho language. However, this claim is controversial as the earliest dates for Homo predate Au. sediba by about 500,000 years (Becoming Human c2008).
Au. sediba shares some traits with Au. africanus, e.g., cranial capacity, brow ridges, and enlarged tooth cusps (the pointed tops of teeth), causing some to claim the two species are related. Its arm-to-leg proportions are likewise similar to the australopiths. The features that link Au. sediba to Homo include the pelvis shape, more vertical brain case, smaller cheek bones, and molar shape.
Another controversy recently emerged in 2014 when Ella Been and Yoel Rak presented their spinal cord study (Barras 2014). They claim that the spinal cord features and the jawbones of the two specimens identified by Berger are actually two different species: early Homo and an australopith. Additionally, Been and Rak claim the Malapa collection contains the remains of four individuals, not two. Berger disputes this claim that there are two species present. You can read more details about this debate atNew Scientist [optional].
Robust Australopiths
Note: There is debate over whether the differences mentioned below qualify the robust australopiths to be in a separate genus from the australopithecines. In some anthropological works the genus Australopithecus is used. In others, such as this work, Paranthropus is used.
Three robust species of hominins emerged in the Plio-Pleistocene period: Paranthropus aethiopicus, Paranthropus boisei, and Paranthropus robustus. They have morphological features that suggest they were well adapted for eating hard foods that needed grinding, which led to their being identified as “robust.” Their incisors and canines are small, while their premolars and molars are quite large. The zygomatic arch is flared, allowing for larger masticatory muscles that are needed to grind hard foods. This tends to make the face dish-like. A prominent sagittal crest tops the skull, yet another chewing adaptation as it the hard-working temporalis muscle a sturdy bone on which to attach. It has been argued that the robust australopiths were so well adapted that they could not respond quickly enough to changing environmental conditions, which led to their extinction by 1.0mya. Recently, dental evidence from P. boisei suggests that hard foods were not the primary diet of at least P. boisei, but were fall back foods, or foods that were ate when preferred foods were not available (Becoming Human c2008).
Paranthropus aethiopicus

Paranthropus aethiopicus is dated 2.7-2.3mya. While it was first identified as a species in 1967 by French researchers who found a toothless mandible at Omo, Ethiopia, it was not accepted until 1985 when Alan Walker and Richard Leakey found the “black skull” at Lake Turkana, Kenya. It is called the black skull because during the fossilization process it was stained black by minerals present in the sediment. Other P. aethiopicus fossils have been found at Laetoli, Kenya since the 1985 discovery; however, no postcranial material has ever been recovered.
P. aethiopicus exhibits a powerful jaw, well-developed sagittal crest, and megadont, or extremely large, molars. Its cranial capacity, the joints between the jaw and cranium, and prognathic face are similar to Au. afarensis. The prominence of the sagittal crest toward the back of the skull is also similar to Au. afarensis. These features are not present in other robust australopiths or Au. africanus. These similarities lead some paleoanthropologists to claim Au. afarensis is ancestral to P. aethiopicus.
P. aethiopicus shares several characteristics with other robust species such as forward positioned zygomatic arches, a dished face (see P. boisei reconstruction below), thick palate bones, and large premolars and molars.
Paranthropus boisei
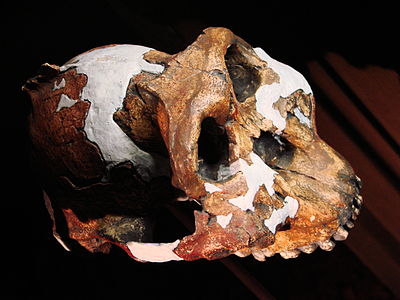
Paranthropus boisei shares several features with P. aethiopicus, including a dished face, strong sagittal crest, and large premolars and molars (in fact the molars are about four times as large as modern humans (Smithsonian 2015)). Unlike P. aethiopicus, the sagittal crest is positioned towards the front of the skull instead of the back. Other unique features include premolars shaped like molars, the zygomatic arch forms a circular arch, the foramen magnum is short and heart-shaped, and the braincase is large than the other robust australopiths. Little was known about the postcranial structures of P. boisei until a 2010/11 discovery of fragments of postcranial remains at Olduvai Gorge, Tanzania. Fragments of the radius, ulna, femur, and tibia support earlier suggestions that P. boisei was sexually dimorphic. These postcranial remains indicate that P. boisei was “…more ruggedly built—combining terrestrial bipedal locomotion and some arboreal behaviors…it [has] more well-formed forearms muscles that were used for climbing, fine-manipulation and all sorts of behavior” (Sci-News.com 2013).
P. boisei was discovered in 1955 at Olduvai Gorge by Mary Leakey. It was not until 1959 that Leakey identified the fossil as a new species and it Zinjanthropus boisei, nicknamed “Zinj.” Subsequent discoveries of similar fossils and other robust australopiths resulted in a consensus among researchers that Zinj belonged in the genus Paranthropus. P. boisei fossils have been found in several East Africa sites, including Peninj, Tanzania, Konso, Ethiopia, and Koobi Fora and West Turkana, both in Kenya. It lived in grasslands, but also forested areas near water sources, between 2.3-1.2mya, making it a long-lived species. There is no evidence to suggest that P. boisei is ancestral to any subsequent hominin.
Paranthropus robustus
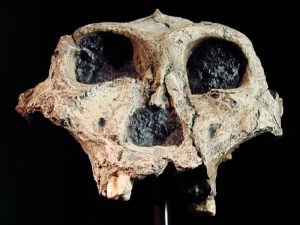
Paranthropus robustus is the only robust australopith found in southern Africa. First discovered in 1938 by Robert Broom in Kromdraii, South Africa, P. robustus fossils were also recovered from Swartkrans and Drimolen, South Africa. Like the other robust species, it has megadont cheek teeth (premolars and molars), large zygomatic arches, dish-shaped face, sagittal crest, and a high degree of sexual dimorphism. Its unique characteristics include a larger brain size than other australopiths and hand morphology suggesting a grip capable of tool use. P. robustus is dated 2.0-1.2mya.
References
Alemseged Z, Spoor F, Kimbel WH, Bobe R, Geraads D, Reed D, Wynn JG. 2006. A juvenile early hominin skeleton from Dikika, Ethiopoa. Nature 443 (Sep 21): 296-301. Available from: http://www.nature.com.offcampus.lib.washington.edu/nature/journal/v443/n7109/full/nature05047.html. doi:10.1038/nature05047.
Bailey SE. 2006. Sahelanthropus tchadensis. In: Encyclopedia of anthropology, Vol. 5. Thousand Oaks (CA): SAGE Reference. p. 2044-2045.
Barras C. 2014. Human ‘missing link’ fossils may be a jumble of species. New Sci [Internet] [cited 2015 Aug 13]; 222(2964). Available from:https://www.newscientist.com/article/mg22229643-200-human-missing-link-fossils-may-be-jumble-of-species/
Becoming Human. c2008. The human lineage through time. Institute of Human Origins [Internet] [cited 2015 Aug 3]. Available from:http://www.becominghuman.org/node/human-lineage-through-time
Domínguez-Rodrigo M, Pickering TR, Baquedano E, Mabulla A, Mark DF, Musiba C, et al. 2013. First partial skeleton of a 1.34-million-year-old Paranthropus boisei from Bed II, Olduvai Gorge, Tanzania. PLoS ONE 8(12): e80347. Available from: http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0080347. doi:10.1371/journal.pone.0080347
eFossils [Internet] [cited 2015 Aug 10]. Department of Anthropology, The University of Texas at Austin. Available from: http://efossils.org/
Harmand S, Lewis JE, Feibel CS, Lepre CJ, Prat S, Lenoble A, Boës X, Quinn RL, Brenet M, Arroyo A, Taylor N, Clément S, Daver G, Brugal JP, Leakey L, Mortlock RA, Wright JD, Lokorodi S, Kirwa C, Kent DV, Roche H. 2015. 3.3-million-year-old tools from Lomekwi 3, West Turkana, Kenya. Nature 521 (May 21): 310-315. Available from: http://www.nature.com.offcampus.lib.washington.edu/nature/journal/v521/n7552/full/nature14464.html. doi:10.1038/nature14464.
Hunt KD. Australopithecines. In: Encyclopedia of anthropology, Vol. 1. Thousand Oaks (CA): SAGE Reference, 2006. p.311-317.
Jurmain R, Kilgore L. Trevathan W. 2013 Essentials in physical anthropology. Belmont (CA): Wadsworth Cengage Learning.
Larsen, CS. 2014. Our origins: discovering physical anthropology. New York (NY): W. W. Norton & Company, Inc.
Lebatard AE, Bouriès DL, Duriner P, Jolivet M, Braucjer R, Carcaillet J, Schuster M, Arnaud N, Monié P, Lihoreau F, Likius A, Macaye HT, Vignaud P, Brunet M. 2008. Cosmogenic nuclide dating of Sahelanthropus tchadensis and Australopithecus bahrelghazali: Mio-Pliecene hominids from Chad. Proc Natl Acad Sci U S A [Internet] [cited 2015 Aug 3]; 105(9): 3226-3231. Available from: http://www.pnas.org/content/105/9/3226.full. doi: 10.1073/pnas.0708015105
Richmond BG, Jungers WL. 2008. Orrorin tugenensis femoral morphology and the evolution of hominin bipedalism. Science [Internet] [cited 2015 Aug 3]; 319(5870): 1662-1665. Available from: http://www.jstor.org/stable/20053635
Scarre C. 2013. The human past. London (UK): Thames & Hudson.
Sci-News.com [Internet]. 2013 Dec 6. Paranthropus boisei: 1.34-million-year-old hominin found in Tanzania. [cited 2015 Aug 12]. Available from: http://www.sci-news.com/othersciences/anthropology/science-paranthropus-boisei-hominin-tanzania-01603.html
Smithsonian Institution [Internet]. 2015 Aug 4. What does it mean to be human? [cited 2015 Aug 11]. Available from: http://humanorigins.si.edu/evidence/human-fossils/species
Su DF. 2013. The earliest hominins: Sahelanthropus, Orrorin, and Ardipithecus. The Nature Education Knowledge Project [Internet] [cited 2015 Aug 3]. Available from: http://www.nature.com/scitable/knowledge/library/the-earliest-hominins-sahelanthropus-orrorin-and-ardipithecus-67648286
Ward CV, Manthi FK, Plavcan JM. 2013. New fossils of Australopithecus anamensis from Kanapoi, West Turkana, Kenya (2003-2008). J Hum Evol 65(5): 501-524. Available from doi:10.1016/j.jhevol.2013.05.006
Wong K. 2006. Special report: Lucy’s baby: an extraordinary new human fossil comes to light. Sci Am [Internet] [cited 2015 Aug 10]; Sep 20. Available from: http://www.scientificamerican.com/article/special-report-lucys-baby/
Cite this page
APA Style: Saneda, T. M. 2022. Proto- and early hominins. In T. M. Saneda & M. Field, Biological Anthropology: a brief introduction. Cascadia College Pressbooks.
Chicago Style: Saneda, Tori M. 2022. “Proto- and Early Hominins.” In Biological Anthropology: A Brief Introduction, 3rd. Bothell, WA: Cascadia College Pressbooks.
CSE Style: Saneda TM. 2010. Proto- and early hominins. In: Biological Anthropology: a brief introduction, 3rd ed. Bothell (WA): Cascadia College Pressbooks. [modified 2022; accessed 2022 Dec 5]. https://openwa.pressbooks.pub/anth205bioanth/chapter/early-hominins.

{kind=link}
{kind=link}
{kind=link}
,_4,4_milioni_di_anni_fa_01.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.JPG){kind=link}
.JPG){kind=link}
.JPG){kind=link}