5 How Did the Hominins Evolve?
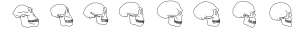
The term hominin is used to differentiate humans and their ancestors from other apes. Hominins share some derived traits, including habitual walking on two legs or bipedalism, big (compared to body size) and more complex brains, and a non-honing chewing complex. As hominins evolve, we see the evolution of not only morphological characteristics but cultural characteristics as well. We can sum up the trends of human evolution as:
- Bipedalism (7.0 mya)
- Non-honing chewing complex (5.5 mya)
- Material culture – stone tools (3.3 mya)
- Larger complex brains (2.8 mya)
- Hunting (1.0 mya).
As you can see from the dates posted, not all these characteristics arose at the same time. In fact, we see variety in when the traits appear in the various hominin paleospecies. There is even variety in the development of the individual trends. For instance, we see evidence of bipedalism about 7.0 mya but we do not see fully bipedal hominins until about 4.2 mya.
Trends in Human Evolution
Bipedalism
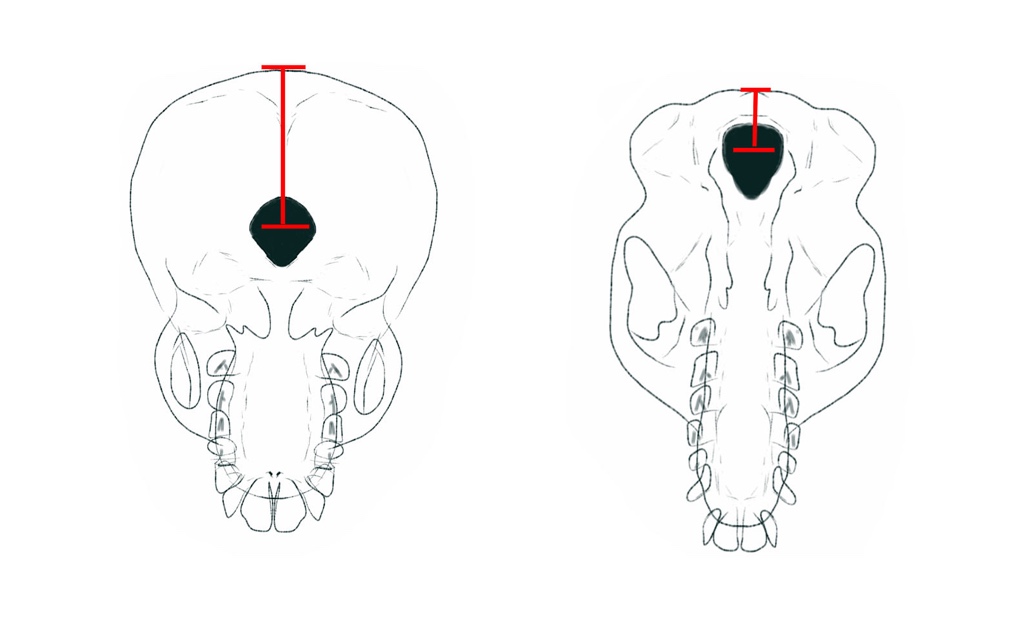
Bipedalism is the first trait to emerge on the road from the ape ancestor to antomically modern humans (AMH). Oddly, the first evidence of habitual bipedalism is not found in the leg structure, but in the skull structure of the proto-hominin, Sahelanthropus tchadensis. Proto-hominins are most easily thought of as transitional species between the ape ancestor and the first fully bipedal hominins. They show evidence of structural changes associated with efficient bipedalism. There is debate among paleoanthropologists as to the degree of bipedalism the proto-hominins displayed. For the earliest, S. tchadensis (7.0-6.0 mya), we can see that the foramen magnum, the hole at the base of the skull where the spinal cord enters the brain case, is more forward and under the skull than at the back of the skull, which is where it is found on apes and other quadrupedal animals. In the graphic below, you can see the position of the foramen magnum on a chimpanzee skull (right) and a human skull (left). The position of the foramen magnum is important for efficient bipedalism. The spinal cord enters under the skull allowing for the forward facing of the eyes while upright.
In subsequent hominin paleospecies, we see that the halluces (big toes) come in line with the other toes, allowing for better balance and power when taking a step forward, an s-curve develops in the spine lowering the center of gravity, the pelvis becomes bowl-shaped creating different attachments of the leg muscles and femur to allow for the forward movement of the leg, as well as the emergence of a padded heel and an arch in the foot both of which provide shock absorption. These are a few of the many morphological changes that evolved for efficient bipedal locomotion. You can explore more about the evolution of bipedalism at eFossils.
One question that paleoanthropologists have explored is why bipedalism evolved in hominins. What evolutionary advantages does it provide? No one answer is adequate to explain its evolution; however, there are multiple advantages it provides (Welsch et. al. 2017, 211):
- It is a more energy efficient method of moving around for long distances, which would have been beneficial as the landscape changed from the tropical rain forests to open savanna. Eventually, the long-distance efficiency would be beneficial when hominins learned to hunt.
- It aids in predator avoidance as bipeds would have been able to see over the savanna grasses.
- It allows for enhanced ability to reach for hanging fruits and plants.
- It decreases the amount of surface area exposed to the sun, thereby providing better heat regulation, which would have been beneficial as the environment changed.
- It frees up hands to carry things
While it is relatively easy to identify the advantages of bipedalism, none can fully explain the evolution of bipedalism on its own, especially when the first bipedal hominins have been found in forested regions of sub-Saharan Africa. It is likely that it was the accumulation of small morphological changes that provided small benefits that ended in efficient habitual bipedalism.
Non-honing Chewing Complex
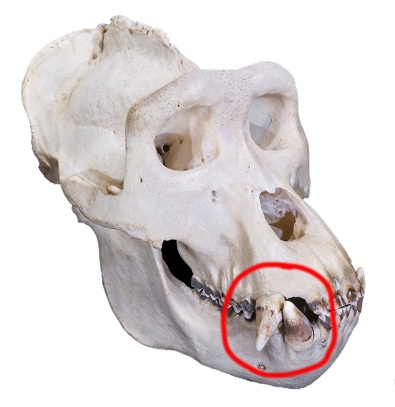
The way the teeth of hominins and their ape cousins process food differs. Apes with the honing chewing complex grab food with their front teeth, using the upper canines and lower third premolars to cut and shred. The upper canines have a sharp back that rubs against the front of the lower third premolar, sharpening both as the jaws open and close (Larsen 2022, 280-281). Hominins lose the honing complex early in their evolutionary trajectory as evidenced by the loss of the gap, or diastema, between the incisors and the canines and the reduction in size of the canines, which become blunt and nonprojecting (Larsen 2022, 281).
Larger, More Complex Brains
“The human brain, with its unequaled cognitive capacity, evolved rapidly and dramatically” (University of Colorado Denver 2012). We see relatively small changes in brain size from the proto-hominins to the early hominins, such as, Australopithecus and Paranthropus. It is not until early Homo (2.8 mya) that we find not only significantly larger brains but more complex brains. The rewiring or neurological reordering of the brain created the basis for changes in cognitive function and behavioral patterns. There is evidence that this rewiring also occurred in later australopithecine paleospecies such as Australopithecus sediba (2.0 mya) (Bryn 2011) but was advanced in genus Homo. Several explanations have been proposed for the expansion of the hominin brain. Dumas et.al. (2012) suggest having more copies of the protein domain DUF1220 resulted in the larger hominin brain. Another hypothesis suggests that “the cognitive challenge associated with social comparison has contributed to cerebral expansion and the disproportionate human brain size…” (Whitaker et.al. 2016). Girskis et.al.’s (2021) study indicates that there was no key gene responsible for the expansion of the hominin brain but was the result of “dozens or perhaps even hundreds” of genes, although Dennis et.al. (2012) claim that duplication of the SRGAP2 gene “…could have allowed for radical—potentially earth-shattering—changes in brain development and brain function.” Whichever turns out to be the case, these changes occurred rapidly and was a special event (Howard Hughes Medical Institute 2005).
Key areas of the brain that changed were the cerebellum, limbic system, and cerebral cortex. The cerebellum functions to coordinate movement. It is what allows us to move smoothly. Recent research suggests that changes in the cerebellum contributed to human culture, language, and tool use (Guevara et.al. 2021). The cerebral cortex controls voluntary movement, language, reasoning, perception, thought, and sensory integration (Szymik 2011). The limbic system is comprised of three structures: the amygdala, hypothalamus, and the thalamus. Together, these structures control our emotions and motivations and are involved in memory (Bailey 2018).
Together, the areas of the brain that were rewired allowed our ancestors to develop cultural responses to changing circumstances, both in the natural environment and in social interactions, including the development of increasingly complex stone tools and sophisticated hunting behavior.
Our Hominin Ancestors
In addition to the trends mentioned previously, as hominins evolve, we will see a shift in morphological characteristics best suited for life in the trees to life on the ground. Over time, hominins lose their curved fingers, arms shorten while legs lengthen. Canines get smaller, molars get bigger, providing the basis for a more omnivorous diet. Hominins proliferate in the changing Pliocene-Pleistocene environments, until we reach the late Pleistocene, when the only hominin left is AMH.
Identifying how many hominin paleospecies have existed is contentious. Not all paleoanthropologists agree on the range of variation we should expect to see in the fossil record, nor can we use reproductive behavior as a factor to identify species because we cannot observe it directly. Using comparative anatomy, paleoanthropologists who expect a wider range of variation lump fossils into fewer paleospecies. These paleoanthropologists are called ‘lumpers.’ Those researchers who expect to see smaller ranges of variation split fossils into more species, thus are called ‘splitters.’ In this chapter, we will take a lumper approach. The chapter will not cover every hominin species, as it is beyond the scope of the introductory anthropology class.
From a student’s perspective, all this complexity probably seems frustrating, it would be ideal if the human story were a straightforward, sequential narrative. Unfortunately, it seems that human evolution was not a nice, neat trajectory of increasingly human-like traits and behaviors; rather it is emblematic of the untidy but exciting nature of the study of human evolution (Yoshida-Levine 2019).
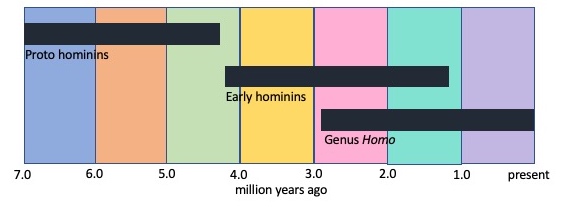
Proto-Hominins
There is debate surrounding the earliest possible members of the hominins. The main issue is that there are not a lot of fossil specimens from this time frame, about 7.0 to 4.4 mya (late Miocene to early Pliocene). Older fossils are simply rarer and the closer we get to the common ancestor of hominins and other apes, the more ape-like it is going to look, making it difficult to differentiate between paleospecies. We do know that we should expect to see a mix of ape-like and human-like features, with the younger fossils displaying more and more human-like features.
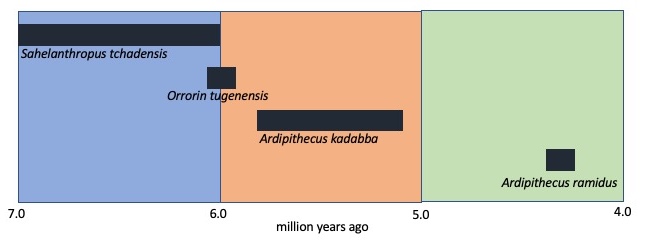
Three genera have been identified as proto-hominins: Sahelanthropus, Orrorin, and Ardipithecus. Some paleoanthropologists claim that these genera are closer to gorillas and chimpanzees then they are to humans. Some have referred to them as ‘hominin imposters’ and that they evolved hominin-like traits in parallel lineages. Others claim that they should all be lumped into the genus Ardipithecus. Sahelanthropus and Orrorin display some evidence of bipedalism, but not much, e.g., position of the foramen magnum for Sahelanthropus and Ardipithecus, and the distribution of the outer bone (cortical bone) in the femur of Orrorin. Based on this limited data, we cannot claim that these hominins were habitual bipeds, only that changes were afoot (forgive the pun).
Early Hominins
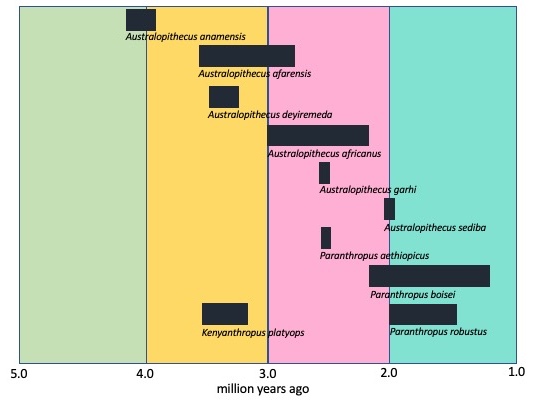
Early hominins span the Pliocene and Pleistocene from 4.2 mya to 1.3 mya and appear to show adaptations to the expanding open savanna environment. One to three genera have been identified. Some researchers lump all of the early hominins into one genus, Australopithecus. Some identify two, Australopithecus and Paranthropus. Others identify three, adding Kenyanthropus. Again, this depends on how the researcher defines the range of variation. Future research will help paleoanthropologists refine what we know about these early hominins. Let’s now explore the three genera in this section.
No matter the genera, the early hominins share some traits:
- They are all fully bipedal. This means that they are habitual bipeds.
- They display intermediate cranial and dental characteristics.
- Ape-like traits:
- prominent jutting forward of the bottom part of the face
- relatively large, forward-slanting incisors
- some still have a diastema
- larger brains than chimpanzees but significantly smaller than modern humans
- curved fingers
- most have longer arms than legs
- flattened foreheads
- Human-like traits:
- smaller canines
- bigger molars
- starting to see a rounding of the skull
- position of foramen magnum
- non-opposable halluces
- leg and pelvic bones display traits associated with bipedalism
- They ate a wide variety of foods, including hard to process foods such as nuts, seeds, roots, and tubers.
- They displayed sexual dimorphism, which means males and females had obvious differences, e.g., size.
- They matured rapidly like chimpanzees, reaching sexual maturity around age four.
Several paleospecies of Australopithecus have been identified: Au. anamensis, Au. afarensis, Au. deyiremeda, and Au. garhi all found in eastern Africa, and Au. africanus and Au. sediba from southern Africa. The East African paleospecies are best known from fossils recovered in Ethiopia, Kenya, and Tanzania, dating from 4.2 mya to 2.5 mya. The paleospecies from southern Africa were found in South Africa, spanning 3.0 mya to 2.0 mya. These early hominins occupied sub-Saharan Africa during the period that the savannas were expanding. Data indicates that might have spent as much time in the trees as they did on the ground. They most likely ate a mixed diet of fruits and leaves (Park 2014). If we were to observe them, we would probably think they looked like walking chimps instead of bipedal humans.
One fossil from this time period that has caused debate is Kenyanthropus platyops. Only one specimen has been found. The team that found it placed it into its own genera due to the unique shape of the face—it is flattened and somewhat dish-like compared to other australopiths. Some argued that the skull had been distorted during the fossilization process and that the skull falls within the range of variation expected for Au. afarensis. Further study using principle component analysis suggested that the specimen fell outside of the range of variation for Au. afarensis, therefore the attribution to a separate genus was warranted (Balter 2009). The debate has not been settled as of the date of this publication and you will see some publications and institutions, e.g., museums and universities, categorize it as Au. afarensis while others have it as a separate genus, Kenyanthropus.
The Paranthropus species, P. aethiopicus, P. boisei, and P. robustus, are often lumped in with the genus Australopithecus primarily because post-cranially they are like the australopithecines. However, others separate them out for their dietary adaptations. All three species are well adapted for heavy chewing as evidenced by their large jaws, flared zygomatic arches (wide bone that flares out behind the cheek bone), massive molars, and sagittal crest (bony protrusion on the top of the skull). The flared zygomatic arch and the sagittal crest are evidence of large facial muscles involved with chewing (temporalis and masseter muscles). Recent microwear analysis and isotopic studies of Paranthropus teeth indicate they were eating low-quality vegetation, e.g., savanna grasses, that required significant grinding by the molars to process (Larsen 2022). Because of the heavy skull features, these species are often referred to as the ‘robust species.’

Genus Homo
The origins of genus Homo are somewhat murky as the Homo fossils from 2.8 to 2.0 mya are fragmentary and poorly preserved. There is also much variability in “brain size, facial features, and teeth and body size” (Yashida-Levine 2019), leading some paleoanthropologists to suggest that any Homo species dating from [2.8] to ~1.6 mya should be excluded from the genus (Tatterall and Schwartz 2009). Others claim that the differences are significant enough to warrant their inclusion. While this might create doubt for some about human evolution, please recall the previous discussion about the range of variation—what should we expect? Scientists know that we should see a mix of characteristics in the transition or early forms of the genus. The AMH skeletal form did not appear overnight—it took millions of years to evolve from its australopithecine ancestor. For this section, you will learn about the prevailing interpretation that the fossils from 2.8 to 1.6 mya are members of genus Homo.
Features that unite the genus include (Park 2014; Yoshida-Levine 2019):
- Smaller, flatter face,
- Smaller jaws and teeth,
- Larger body size,
- Bigger, more complex brains than earlier hominin species,
- Prolonged childhood, and
- Increasing reliance on culture, such as stone tools and cooperative childcare.
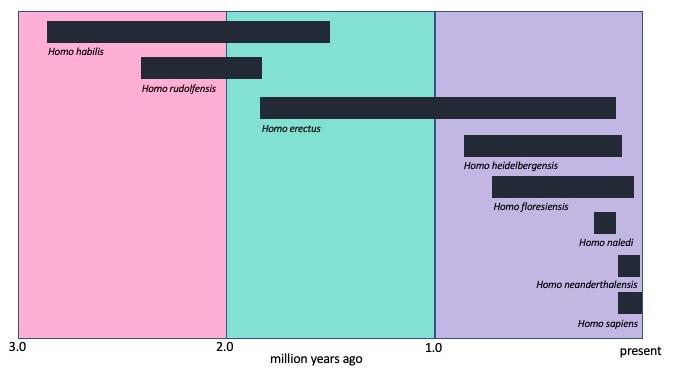
Early Genus Homo
The earliest identified members of the genus are Homo habilis (2.8 mya to 1.5 mya) and Homo rudolfensis (2.4 my to 1.8 mya). While some paleoanthropologists classify these two species as one, H. habilis, claiming that the differences are the result of sexual dimorphism, the differences in the specimens seem to warrant the split into two species. H. habilis has a u-shaped dental arcade while H. rudolfensis has a parabolic arch (click here to see a graphic comparing dental arcades; scroll down until you see “Dental Arches”). H. rudolfensis also has a broad, flat face, larger brain, and a slightly developed brow ridge in comparison to the small, more prognathic face and strongly developed brow ridge of H. habilis. Both lived in the savanna grasslands and were most likely omnivores.
Paleoanthropologists know more about Homo erectus, although some differentiate between fossil specimens in Africa (Homo ergaster) and those in Asia (H. erectus). Asian fossil specimens tend to have a more robust skull and those in western Eurasia have smaller brains and bodies (Yashida-Levine 2019). We will treat it as one species, H. erectus. One of the significant changes from earlier Homo species is the increase in brain size. The brain size of H. habilis averaged about 650 cubic centimeters (cc) while H. erectus averaged 950 cc. For the first time, hominins approximate modern human height (5’ 4”) and have a fully modern bipedal gait. If you saw a H. erectus walking down the street, you would not be able to differentiate their ability to walk from ours, although its head would give it away as not-quite-human. The cranial features are more robust than AMH with a heavy brow ridge and a more elongated head. At the back of the skull, you would see a horizontal ridge called an occipital torus and a slight bony ridge on top called a sagittal keel.
From one of the statements above, you should be able to surmise that this is the first hominin to leave Africa. In fact, this species has been recovered in Africa, Asia, and Europe. Fossils dated to 1.8 mya have been found in Africa, Eurasia, and Indonesia, where the first H. erectus fossil was discovered in Java in 1891. H. erectus appears in the European fossil record by 1.2 mya and in China by 700 thousand years ago (kya), although Zhu et.al. (2015) claim H. erectus was in China as early as 1.6 mya. It disappears from the fossil record in Asia and Africa by 300 kya and in Europe by 400 kya.
Hopefully, you are asking yourself, “How did H. erectus survive in such a wide range of environments?” The answer is cultural adaptations. We know definitively that all the Homo species could manufacture and use stone tools and most likely some of the later australopiths could as well. However, australopithecine fossils have not been found associated with stone tools, but stone tools have been found in stratigraphic layers dated to the time australopiths lived. The Lomekwi tool industry is dated to 3.3 mya. Found at the Lomekwi 3 site in West Turkana, Kenya, these simple stone tools represent a cognitive evolution for early hominins. The ability to knap even simple stone tools requires the ability to understand the properties of stone—which stone when knapped will create stone flakes and which will not. The only hominin that has been found in the area and dated to this time is K. platyops.

The next oldest tool industry is the Oldowan tool industry, first found at Olduvai Gorge, Tanzania and dated to 2.6 mya at Gona, Ethiopia. Oldowan tools were found associated with H. habilis. In fact, it was this association that gave rise to the name of the species, which translates to ‘handy man.’ Both Lomewki and Oldowan tool industries used hard percussive manufacturing techniques, meaning that two stones were struck together to create the tools. The stone tools associated primarily with H. erectus used a soft percussive technique that required an intermediary material such as an antler or a piece of bone to be used between the two stones, allowing the knapper to have more control and create sharper edges. This stone tool industry is called the Acheulean.

The Acheulean tool industry, used from 1.76 mya to 200 kya, represents another cognitive leap for hominins. In fact, some researchers suggest that the ability to make Acheulean tools was genetically hardwired in H. erectus and Homo heidelbergensis (Hirst 2019). These bifacial (meaning worked on both sides) stone tools required the maker to mentally visualize the finished product while understanding the breakage properties of the stone. The tool industry is characterized by the hand axe pictured above. This multi-purpose tool has often been found associated with the remains of large animals, inferring that the tools were used to kill and butcher the animals (Larsen 2022, 336). Incorporating more meat into their diets was one way H. erectus could have supported the metabolic needs of their large brains. This ability to hunt may have been one of the behaviors that allowed them to successfully inhabit a wide variety of environments. The ability to at least harness the use of fire would be another behavior that enabled survival in the colder, glacial conditions of the northern hemisphere.
The evidence that H. erectus could use fire comes from several sites, including Koobi Fora, Kenya (1.5 mya), Wonderwerk Cave, South Africa (1.0 mya) and Zhoukoudian, China (300 kya). At the latter site, burned animal bones, seeds, and stone tools suggest that H. erectus was cooking food. Additionally, fire would have provided warmth and protection.
Hominins’ increasing reliance on tools profoundly affected human biology. As tools began to perform the functions of the face and jaws in preparing food for consumption—that is, in cutting up, cooking, and processing meat and other food—there was a commensurate decline in the robusticity of body parts associated with mastication. In terms of both culture and biology, H. erectus evolved the contextual behavior—hunting, successful dispersal across large territory, adaptive success, and increasing dependence on and effective use of culture–as a means of survival. The increased dependence on culture and the dominance of behaviors requiring technology for the acquisition and processing of food increased the diversity of environments occupied by H. erectus (Larsen 2022, 341).
Interestingly, no evidence of cultural behavior (other than the deposition of the remains themselves) has been found associated with Homo naledi, a fossil hominin with cranial features similar to H. erectus. These similar features, large brow ridge, sagittal keel, small teeth, and thick cranial bones, were not enough to offset the dissimilarities, including the australopith-like features such as small brain size, curved fingers, and body size. The large number of fossils found at the Rising Star Cave site, South Africa, is the largest deposit of hominin remains found at any one site in Africa with over 1,300 fossils representing at least a dozen individuals (Berger and Hawks 2017). H. naledi‘s human-like features, including a foot “nearly indistinguishable” from an AMH foot, hand and wrist bones, and vertebra, cinched that the specimens represented a new paleospecies dating to 335 kya to 236 kya (Dirks et.al., 2017). The main question Berger and his team are still trying to answer is why a relatively primitive Homo species existed at a time of more advanced Homo species. You can listen to Berger describe the finding and excavation of the Rising Star Cave on YouTube. Not only is the excavation known for the discovery of a new hominin species, but it is the first all-female excavation team assembled to investigate a paleoanthropological site.
Late Genus Homo
One of those more advanced species was H. heidelbergensis, dating from 800 kya to 130 kya. Some paleoanthropologists include H. heidelbergensis in a generic group called archaic H. sapiens with other species including Homo antecessor, Homo georgicus, Homo rhodesiensis, and H. neanderthalensis. In this text, H. neanderthalensis is treated as a distinct species, while H. antecessor, H. georgicus, and H. rhodesiensis are lumped together with H. heidelbergensis. What unites these species is increasing brain size, average 1200 cc, an emerging forehead, mix of H. erectus-like and H. sapiens-like skull features, slightly smaller teeth and more gracile cranial bones, but robust post-cranial bones; however, there is regional variation in the fossil population—some are more like H. erectus and some are more like H. sapiens (Paskey and Cisneros 2019). Data does suggest that H. erectus is ancestral to H. heidelbergensis, which is ancestral to both H. neanderthalensis and H. sapiens (Buck and Stringer 2014; Rightmire 2013).
It is interesting to note that while overall teeth tended to be smaller than earlier hominins, the incisors became larger. During this time, the front teeth show signs of severe wear. In fact, some are worn down almost to the gum line. Paleoanthropologists interpret this as H. heidelbergensis using their front teeth as a gripping tool (Larsen 2022). Other cultural innovations include the construction of free-standing structures. The best evidence for this comes from Terra Amata, France, where habitation debris was found bound together in what appears to be the floor of a structure. This debris informs us that fishing was a primary occupation at this site, some of the earliest evidence for this subsistence strategy. H. heidelbergensis most likely hunted big game using an ambush strategy that involved forcing the animals off cliffs or cornering them in dead-end canyons as evidenced at sites in the Channel Islands, France (Welker 2017.). Sites like Schöningen, Germany, have provided additional support for hunting. Over ten thousand organic artifacts and ecofacts, including animal bones, were recovered from the site as well as long, wooden spears dating to 400 kya. These javelin-like weapons were preserved in mud underlying a peat layer, both of which are known to inhibit the growth of bacteria that breaks down organic matter. The spears were pointed at both ends and show evidence that they were fire hardened. The structure and weight of the spears indicate that they were made to be thrown and are the oldest evidence of wooden throwing spears in the archaeological record (Thieme 1997).
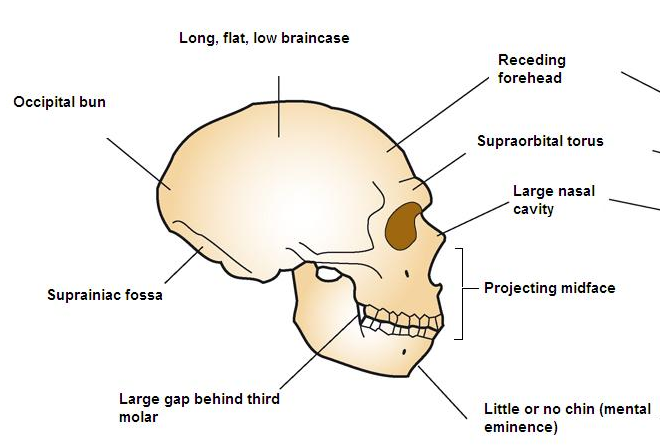
Homo neanderthalensis is the only hominin to have evolved in cold weather. Some of their morphological traits are adapted for the glacial environments in which they evolved, such as the large nasal aperture (the pear-shaped opening in the skull where the fleshy nose would be) that provided ample space for adding moisture to the cold air they were breathing (and warming it as well) and their short, stocky stature. Having a short, stocky form meant that they had less body surface area from which to radiate heat. Other physical characteristics included a projecting mid-face, large brow ridges (supraorbital torus), widely spaced eye orbits, ‘swept-back’ cheek bones, a low, long braincase, and an occipital bun. Neanderthal’s brains were large, on average 1500 cc. The size and shape of the Neanderthal brain suggests that Neanderthals matured more slowly than Homo sapiens (Goldfield 2019).

Proto-Neanderthals, hominins with intermediate traits between H. heidelbergensis and H. neanderthalensis, first appear in the archaeological record about 400 kya at Sima de los Huertos, Atapercua, Spain. Full-fledged Neanderthals appear in the archaeological record at about 200 kya but disappear by 30 kya. The oldest Neanderthal remains are found in Ice Age Europe; however, sites in the Middle East tell us that Neanderthals migrated out of the region during the coldest glacial period. While Neanderthals did evolve physical adaptations to the cold environment, they developed cultural ways to adapt as well. One of these ways was a better tool industry, the Mousterian, to extract resources more easily. Mousterian tools were developed for specific purposes, e.g., scrapers, projectile points, and blades. Which type of tool predominated the tool kit varies by region, which may provide insight into subsistence strategies and other behaviors adapted for specific environments. With their improved tool kit (compared to earlier tool industries), Neanderthals were able to successfully hunt. Isotopic analysis of Neanderthal bones plus the amount of animal bone found at Neanderthal sites indicate that meat, especially that of reindeer and mammoth, dominated the Neanderthal diet. This is unsurprising as the warm months when vegetation would have been available would have been short in the glacial environment.
Neanderthals utilized caves and rock shelters in southern Europe during the coldest parts of the year, then moved to the open-air camps, most likely to follow herd animals north during their migration. Caves appear to have been used long term, suggesting groups returned to these sites over and over, or perhaps part of the group used the cave sites year-round while others hunted herd animals. The presence of post molds at cave and rock shelters informs archaeologists that Neanderthals covered entrances to block cold air. A post mold is created when the wood or bone from a post decays, leaving a dark circle behind. The pattern of the post molds is used to determine the shape and size of structures. Other patterns found at Neanderthal sites indicate that they were building structures to enclose themselves, most likely covering the posts with animal hides. Multiple hearths inside the structures may indicate that multiple family groups inhabited the same structure.
That Neanderthals lived in some type of social group is further supported by the presence of burials. Several sites demonstrate the intentional burial of the dead, including La Chapelle-aux-Saints, France, Amud, Israel, Tabun, Israel, and Shanidar Cave, Iraq. Shanidar is of further importance as evidence of the medicinal use of plants has been found associated with an individual who lived until about the age of 40 (that would be like someone living until they were 90 in US culture!). This individual, a biological male, had survived crippling arthritis and several major wounds, including one that blinded him, to live to a ripe old age. This could only have happened if he had someone to take care of him. It has been argued that burials were simply a way to dispose of dead bodies; however, the bodies seem to be intentionally buried as almost all are found in the fetal position with their hands deliberately placed. It suggests that there was a symbolic element to the burials. Some Neanderthal dead provide evidence of cannibalism. At Krapina, Croatia, El Sidrón, Spain, Moula-Guercy Cave, France, and Goyet, Belgium, the Neanderthal bones show the same type of cut patterns found on animal bone (Larsen 2022). Whether it was for food or ceremonial reason is unknown.
More evidence of Neanderthal symbolic behavior comes in the form of artistic expression. At Cueva de los Aviones and Cueva Antón in Spain, archaeologists found perforated marine shells painted with different colors of ochre. Dating to ~ 115 kya, this is the earliest evidence that Neanderthals ornamented themselves, perhaps to express identity. Evidence that H. sapiens created similar artifacts dates to 120 kya in Africa. Previously, this artistic expression was seen as exclusive to H. sapiens, but this recent research caused researchers to recontextualize their thoughts about Neanderthals. The remains of bird wings at Fumane Cave, Italy (44 kya), and other cave sites in France indicate that feathers were of interest to our hominin cousins (Peresani et.al. 2011). Deep cross-hatching at Gorham’s Cave, Gibraltar (39 kya), adds yet another form of self-expression practiced by Neanderthals (Rodríguez-Vidal et.al., 2014).

It is now clear that Neanderthals engaged in one the most symbolic behaviors—talking. There are three primary lines of evidence:
- Microscopic wear patterns on incisors and canines coupled with what we know about brain laterality (right/left brain; handedness is an indication of a lateral brain, and we know the left side of the brain is instrumental in speech; scratches left on the teeth can indicate whether an individual is right- or left-handed) provide a strong line of evidence that Neanderthals could speak
- The hypoid bone from a Neanderthal skeleton, a critical structure in the ability to speak, was found Kebara, Israel, and,
- DNA from a Neanderthal skeleton at El Sidrón, shows the presence of the FOXP2 gene, a gene important in speech production (Larsen 2022).
Contrary to what early paleoanthropologists claimed about Neanderthals (and what the public generally believe) , that they were unintelligent brutes, modern evidence indicates the exact opposite. Neanderthals had a complex culture, complete with social groups and sophisticated methods of survival. It most likely explains why modern H. sapiens interbred with Neanderthals as long as 65 kya to 50 kya (Raff 2022)—the Neanderthals did not appear all that different from themselves. DNA evidence shows that most modern human populations contain some Neanderthal (1-4%) except for sub-Saharan African populations, indicating that interbreeding occurred, most likely in the Middle East (Slatkin and Racimo 2016). All of which begs the question, why did the Neanderthals disappear from the archaeological record? It is unlikely that there is one reason for this, e.g., no comet hitting Europe wiped them out. Instead, there was a complex interaction of cultural interactions, biological characteristics, and environmental pressures that to their genome being assimilated into AMHs even as culturally, their behaviors were lost.
Neanderthals evolved in isolation from H. sapiens, who did not arrive in Europe until about 54 kya (Slimak et.al., 2022), which is also about the time Neanderthals could have encountered AMHs in the Middle East. About 70-60 kya, a particularly cold period began, the Weichsel glacial period. As temperatures plummeted, the landscape changed. With fewer resources available, Neanderthals began to move south into the Middle East. After this cold period, the climate began to warm again, beginning a series of cooling and warming cycles. It is also possible that a volcanic eruption 40 kya further stressed resources. One hypothesis about why Neanderthals disappeared was that physiologically they could not adapt to these changing environmental conditions because they were too well adapted to the conditions they initially evolved in. At the same time, they did not adapt quickly to changing resources, and could not compete technologically for those resources with AMHs, who had a highly sophisticated tool kit to extract both terrestrial and aquatic resources. Another hypothesis suggests that it is possible that Neanderthals’ cognitive abilities differed from modern humans, which might have put them at a disadvantage in the changing circumstances. And, if we consider the possibility that their maturation cycle was longer then modern humans, then there are multiple factors that could have contributed to their disappearance. And yet another hypothesis suggests that Neanderthal males carried a gene on the Y-chromosome that might have caused a maternal immune response during gestation, negatively impacting reproduction (Mendez et.al., 2016). If Neanderthals had small populations as proposed (anywhere from 5,000 to 70,000 individuals) any stressor or combination of stressors would have been difficult to respond to (Paskey and Cisneros 2019). No matter the causes, Neanderthals were no longer in northwest Europe by 40 kya (Devièse et.al., 2021) and everywhere else by 28 kya (Paskey and Cisneros 2019).
Meanwhile, in the islands of southeast Asia, a small population of genus Homo was adapting to a highly different environment. On the Indonesian island of Flores, researchers found skeletal remains at Liang Bua Cave. Dating to between 74 kya to 12 kya, Homo floresiensis was a small hominin, more the height of an australopith at 3.3 feet with a small brain (400 cc) and large feet. The combination of traits made the popular press dub the skeleton ‘The Hobbit.’ Subsequent skeletal discoveries on the island put its average height at 3.5 feet and weighing anywhere from 35-79 pounds. Regarding its brain size, yes, it is small, but in comparison to its body size, the brain is similar to what we would expect to see in early Homo genera. While the place of the paleospecies within the hominin tree is still debated, most paleoanthropologists think that H. floresiensis is descended from H. erectus. One might raise an eyebrow as how the shortest hominin in genus Homo is descended from the first hominin to reach modern human height. Consider the isolated environment of an island with a finite number of resources. Large individuals may have problems finding enough nourishment to sustain their size. Smaller individuals would have less trouble, thereby giving them an evolutionary advantage over their larger brethren. This process of decreasing size has been documented in other species like the dwarf stegodon, which is a relative of Asian elephants, and is called island dwarfism or insular dwarfism. While a few researchers suggest H. floresiensis suffered from microcephaly, a disorder that causes a small brain case and limited cognitive abilities, the stone tools that have been found do not support this possibility. Insular dwarfism is the better explanation based on current data. There is no evidence that H. floresiensis interacted with AMH before it became extinct around 12 kya, possibly due to volcanic activity (Paskey and Cisneros 2019).
In 2010, scientists announced the discovery and DNA analysis of a finger bone and two teeth found in Denisova Cave, Siberia (Reich et al. 2010). The skeletal remains were recovered from a deposit dated to 50 kya to 30 kya. More recent discoveries indicate Denisovans were in Asia at least 160 kya (Chen et.al. 2019; Demeter et.al. 2022). Data suggest that the remains were from an individual who shared a common origin with Neanderthals, but was not a Neanderthal, nor was it a modern human. It was from a group that split from Neanderthals after AMH split from H. heidelbergensis (Slotkin and Racimo 2016). However, DNA evidence tells us that the Denisovans share 4-6% of its genetic material with modern peoples living in New Guinea, Bougainville Islands, and China. Further studies (Cooper and Stringer 2013) indicate that the Denisovans crossed Eurasia and interbred with modern humans about 55 kya to 45 kya (Raff 2022), but still, genetically the Denisovans were more closely related to Neanderthals then modern humans (Meyer et al. 2012). While further finds will shed more light on the Denisovans, there was more genetic variability during the Pleistocene than previously thought (Larsen 2014), and gene flow between the various populations has implications for the emergence of modern humans (Pääbo 2015).
Anatomically Modern Humans
At present, only one hominin species inhabits planet Earth, us, H. sapiens. Morphologically, H. sapiens characteristics include the presence of a chin (or mental prominence), a large brain, flat face, rounded or globular cranium, and a continuous, reduced brow ridge. Our bones are gracile in comparison to earlier hominins, although the earliest anatomically modern H. sapiens are more robust than we are today and shows none of the cold weather adaptations of Neanderthals. The more gracile nature lends credence to the hypothesis that AMH evolved first in Africa. The leaner body proportions are more adaptive to the tropical African environments as there would be more body surface area to radiate heat. DNA evidence also supports an African origin. Modern African populations have more genetic diversity than any other modern human population, inferring that that they have been evolving longer (Saneda and Field 2015).
The oldest H. sapiens fossils are from North Africa at Jebel Irhoud, Morocco (Richter et. al. 2017). Dated to about 300 kya, the hominin fossils from Jebel Irhoud require a rethinking of the evolution of H, sapiens. Mounier and Lahr (2019) suggest “…Homo sapiens…originated from the coalescence of South and, possibly East-African source populations, while North-African fossils may represent a population which [mated with] Neanderthals during the [Late Middle Pleistocene].” Other African H. sapiens sites include Omo Kibish, Ethiopia, dated to 195 kya, Herto, Ethiopia (160 kya–154 kya), and Klasies River Mouth and Border Cave, South Africa (120 kya – 89 kya). Additionally, H. sapiens fossils from Skhūl, Israel (130 kya–100 kya), Qafzeh, Israel (120 kya–92 kya), Zhoukoudian and Tianyuan in China and Niah Cave, Borneo, Indonesia date to 40 kya or younger and in Europe dating no earlier than 31 kya (Mladeč, Czech Republic), help us trace the evolutionary history of our species. Modern humans made it to Australia by 55 kya; however, no human remains have been found there dating earlier than 30 kya at Lake Mungo. This data lends little support for the Multiregional Hypothesis, a model developed by Milford Wolpoff, which claims that modern H. sapiens evolved in separate places in the Old World from local archaic H. sapiens populations. This model suggested that gene flow between the populations led to the genetic similarity of modern humans. Most anthropologists support the model of an African origin. There are two variants of this model. The Out of Africa model, proposed by Chris Stringer, claims that AMH arose in Africa and spread to Europe after 50 kya, replacing non-H. sapiens population with no gene flow. Fred Smith, Anthony Falsetti, and Steven Donnelly proposed the Assimilation model that also claimed modern human first arose in Africa and spread to Europe and Asia (Larsen 2022). The primary difference between the Out of Africa and Assimilation models is that the latter does claim gene flow between H. sapiens and H. neanderthalensis. Considering the recent DNA analyses of Neanderthals and Denisovans discussed above, multiple lines of evidence support the Assimilation model.
With the emergence of AMH, we see an expansion of behaviors first identified with Neanderthals. Although AMH first used the same stone tools as their contemporaries, by 40 kya in Europe, they were making sophisticated tool kits that varied regionally. For instance, the Aurignacian tool industry (40 kya-28 kya) is characterized by blade-based technology (a blade is twice as long as it is wide) using stone, bone, wood, antler, and shell. It also utilized a new manufacturing technique called the punch technique, which involved using a spear and upper body weight, or a hammerstone, or a hammer and chisel to punch blade from a stone core. The Gravettian (29 kya-22 kya) is characterized by backed blades that could be hafted onto wooden handles and shafts. The stone projectile points, associated with megafauna, or big game, remains such as bison, mammoth, horse, and reindeer, indicate that the Gravettian peoples were efficient hunters. In fact, some have referred to them as the Mammoth Hunters. Other well know tool industries from this time period in Europe, the Upper Paleolithic, are the Solutrean (22 kya-18 kya) and the Magdalenian (17 kya-11 kya). The former is restricted to the Iberian Peninsula and utilized a new manufacturing technique, lithic reduction percussion and pressure flaking. Heat treating the stone, allowed the knapper to use an antler or bone to force small flakes off the edges of tools, making them sharper. The latter tool industry is a blade tool industry characterized by denticulated (serrated) microliths (think thumbnail size projectile points and scrapers), and harpoons. The Magdalenian tool industry is associated with the resettlement of Europe after the last glacial maximum (24 kya-18 kya). It is most likely during the Magdalenian that the atlatl or spear thrower was invented as well as the bow and arrow, making hunting more efficient and safer since the hunter did not need to get as close to their prey. Other hunting tools like bolas (stone missiles), boomerangs, and traps are dated to the Upper Paleolithic period. Clearly, our earliest ancestors were able to adapt to a wide variety of environments relatively quickly and out-compete other hominin species living at the time.
Interestingly, it seems that the Neanderthals may have tried to copy the Aurignacian tool industry. The Châtelperronian tool industry (40 kya-34 kya) has only been found associated with Neanderthals during the Upper Paleolithic and is a mix of Mousterian and Aurignacian type tools. It is possible that Neanderthals developed this tool industry on their own, but the timing with the appearance of Aurignacian tools makes it a topic of discussion.
Anatomically modern humans were good hunters as evidenced by animal remains found at sites; however, the appearance of larger settlements with structures that had sunken and stone foundations, wooden posts, often found in areas where there was no wood available, and mammoth tusk (and antlers of other megafauna) indicate that they were less mobile than Neanderthals. Additionally, the structures were often built on tops of hills, which provided a strategic advantage. We find hearths and cooking ovens at AMH habitation sites along with storage pits and stone lamps in the form of flat stones with a depression containing animal fat. Tailored clothing has been found at the Sungir site (22 kya) in Russia. Coupled with bark cloth and woven grass for insulation, the Upper Paleolithic AMH were culturally adapted to the glacial environments of the north (Welker 2017).
AMHs expand beyond the relatively simple art forms of the Neanderthals to create cave art and portable sculptures in forms that are sometimes recognizable to us even if we cannot reliably interpret the symbolism. Animal forms proliferate as do hunting scenes and abstract forms such as lines and dots. Few representations of humans are found in cave art. When they are found, they appear as anthropomorphic figures, half-human, half-animal. More sculptures are in human form than animal as evidenced by the Venus figurines, which may be the first forms of self-representation. Why AMHs created this art is unknown, although it has been proposed that the art represents myths, magic, and ritual. Some may be to communicate information about hunting, thereby performing a more educational purpose. Some of the most well-known sites for Upper Paleolithic Cave art are Lascaux (17 kya) and Chauvet (32-31 kya) in France and Altamira (15 kya) in Spain.


Perhaps it was the long, cold winters that provided time for AMH to create the abundance of art found at Upper Paleolithic sites in Europe, but it was not just in Ice Age Europe that AMH exhibited artistic expression. Cave art depicting warty pigs has been found in Sulawesi, Indonesia dating to about 44 kya (Brumm et.al. 2021). Various types of shell beads from such sites as Skhul, Israel dated to 100 kya (Balton 2006) and Grotte des Pigeons, Morocco dated to 82 kya (Bouzouggar et.al. 2007), as well as 42 kya shell beads from Riwi Cave, Australia (Balme and Morse 2006), and carved deer bones possibly 40 kya from Turobong Cave, South Korea (Portal 2000).
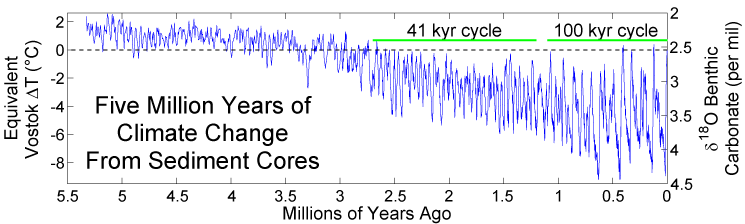
By the end of the Paleolithic period, about 12 kya, AMH inhabited most areas of the world from Australia to Siberia and the Americas and had started the domestication process for some plants, such as fig trees and emmer wheat, and animals, such as European wolves, sheep, and goats. But what drove hominin biological and cultural evolution? The answer, generally speaking, is easy—environmental stressors, both environmental, biological, and cultural. Specifically, it is a complex interaction of these stressors. Climate and environmental change have both been linked to the disappearance and appearance of hominin species. Hunting, patchy forests, and provisioning have been proposed to explain bipedalism, which if you recall is the first morphological trait to appear on the evolutionary road to us. The late Miocene, when we see the first changes associated with bipedalism, was geologically turbulent in East Africa as the terrain was reformed with mountains and the Great African Rift Valley emerged, reducing the amount of forest area. Global cooling in the Pliocene caused the expansion of the polar ice caps and the lowering of sea levels. The Panama land bridge formed causing changes to ocean currents, resulting in weather disruptions (Welker 2017). In Africa, the savannas were expanding while the continent began to experience distinct seasons, e.g., a wet season and a dry season. The Pleistocene, or Ice Age, ushered in an era of global cooling and warming periods, the expansion of glacial sheets of ice over the northern hemisphere, and concomitant rising and lowering of sea levels, all of which impacted animal and plant species. Species adapted or they died out. Hominins would have been competing with each other and other animals for resources. Our earliest ancestors foraged and scavenged for those resources, but over time evolved cultural means to extract the resources they needed, until there was only one hominin species left, us. However, we must continue to adapt to changing circumstances.
References
Almécija, Sergio, Ashley S. Hammond, Nathan E. Thompson, Kelsey D. Pugh, Salvador Moyà-Solà, and David M. Alba. 2021. “Fossil Apes and Human Evolution.” Science 372(6542). Accessed March 22, 2022. https://doi-org.offcampus.lib.washington.edu/10.1242/jeb.240440.
Bailey, Regina. 2018. “The Limbic System of the Brain.” ThoughtCo. Accessed March 28, 2022. https://www.thoughtco.com/limbic-system-anatomy-373200.
Balme, Jane and Kate Morse. 2006. “Shell Beads and Social Behavior in Pleistocene Australia.” Antiquity 80: 799-811. Accessed April 5, 2022. https://www.academia.edu/1996277/Shell_beads_and_social_behaviour_in_Pleistocene_Australia.
Balter, Michael. 2009. “What Ever Happened to Kenyanthropus platyops?” Science. Accessed March 28, 2022. https://www.science.org/content/blog-post/what-ever-happened-kenyanthropus-platyops.
Balton, Michael. 2006. “First Jewelry? Old Shell Beads Suggest Early Use of Symbols.” Science 312(5781): 1731. Accessed April 27, 2022. https://doi-org.offcampus.lib.washington.edu/10.1126/science.312.5781.1731.
Becoming Human. 2008. “The Human Lineage through Time.” Institute of Human Origins. Accessed August 3, 2015. http://www.becominghuman.org/node/human-lineage-through-time.
Berger, Lee and John Hawks. 2017. Almost Human: The Astonishing Tale of Homo naledi and the Discover That Changed Our Human Story. Washington, DC: National Geographic.
Bouzouggar Abdeljalil, Nick Barton, Marian Vanhaeren, Francesco d’Errico, Simon Collcutt, Tom Higham, Edward Hodge, Simon Parfitt, Edward Rhodes, Hean-Luc Schwenninger, Chris Stringer, Elaine Turner, Steven Ward, Abdelkrim Moutmir, and Abdelhamid Stambouli. 2007. “82,000-Year-Old Shell Beads from North Africa and Implications for the Origins of Modern Human Behavior. Proceedings of the National Academy of Sciences of the United States of America 104(24): 9964-9969. Accessed April 27, 2022. https://www.jstor.org/stable/25435867.
Brumm, Adam, Adhi Agus Oktaviana, Basran Burhan, Budianto Hakim, Rustan Lebe, Jian-Xin Zhao, Priyatno Hadi Sulistyarto, Marlon Ririmasse, Shinatria Adhityatama, Iwan Sumantri, and Maxime Aubert. 2021. “Oldest Cave Art Found in Sulawesi.” Science Advances 7(3). Accessed April 5, 2022. https://www.science.org/doi/10.1126/sciadv.abd4648.
Bryn, Brandon. 2011. “Australopithecus sediba May Have Paved the Way for Homo. American Association for the Advancement of Science. Accessed March 28, 2022. https://www.aaas.org/news/science-australopithecus-sediba-may-have-paved-way-homo.
Buck, Laura T. and Chris B. Stringer. 2014. “Homo heidelbergensis.” Cell Biology 24(6): R214-R215. Accessed April 1, 2022. https://doi-org.offcampus.lib.washington.edu/10.1016/j.cub.2013.12.048.
Chen, Fahu, Frido Welker, Chuan-Chou Shen, Shara E. Bailey, Inga Bergmann, Simon Davis, Huan Xia, Hui Wang, Roman Fischer, Sarah E. Friefline, Tsai-Luen Yu, Matthew M. Skinner, Stefanie Stelzer, Guangrong Dong, Qiamei Fu, Guanghui Dong, Jian Wang, Dongju Zhang, and Jean-Jacques Hublin. 2019. “A Middle Pleistocene Denisovan Mandible from the Tibetan Plateau.” Nature 569: 409-412. Accessed May 18, 2022. https://www.nature.com/articles/s41586-019-1139-x.
Cooper, A. and D. B. Stringer. 2013. “Did the Denisovans Cross Wallace’s Line?” Science 342(6156): 321-323. Accessed April 4, 2022. doi:10.1126/science.1244869.
Demeter, Fabrice, Clément Zanolli, Kira E. Westaway, Renoud Joannes-Boyau, Philippe Duringer, Mike W. Morley, Frido Welker, Patrick L. Rüther, Matthew M. Skinner, Hugh McColl, Charleen Gaunitz, Lasse Vinner, Tyler E. Dunn, Jesper V. Olsen, Martin Sikora, Jena-Luc Ponche, Eric Suzzoni, Sébastuen Frangeul, Quentin Boesch, Pierre-Olivier Antoine, Lei Pan, Song Xing, Jian-Xin Zhao, Richard M. Bailey, Souliphance Boualaphane, Phonephanh Sichanthrongtip, Daovee Sihanam, Elise Patole-Edoumba, Françoise Aubaile, Françoise Crozier, Nicolas Bourgon, Alexandra Zachwieja, Thonglith Luangkhoth, Viengkeo Souksavatdy, Thingsa Sayavongkhamdy, Enrico Cappelline, Anne-Marie Bacon, Jean-Jacques Hublin, Eske Willerslev, and Laura Shackelford. 2022. “A Middle Pleistocene Denisovan Molar from the Annamite Chain of Norther Laos.” Nature Communications 13:2557. Accessed May 18, 2022. https://www.nature.com/articles/s41467-022-29923-z.
Dennis Megan Y. Xander Nuttle, Peter H. Sudmant, Francesca Antonacci, Tina A. Graves, Mikhail Nefedov, Jill A. Rosenfeld, Saba Sajjadian, Maika Malig, Holland Kotkiewicz, Cynthia J. Curry, Susan Shafer, Lisa G. Shaffer, Pieter J. de Jong, Richard K. Wilson, and Even E. Eichler. 2012. “Evolution of Human-Specific Neural SRGAP2 Genes by Incomplete Segmental Duplication.” Cell 149(4): P912-P922. Accessed March 28, 2022. https://doi.org/10.1016/j.cell.2012.03.033.
Devièse, Thibaut, Grégory Abrams, Mateja Hajdinjak, Stéphane Pirson, Isabella De Groote, Kévin Di Modica, Michel Toussaint, Valentin Fischer, Dan Comeskey, Luke Spindler, Matthias Meyer, Patrick Semal, and Tom Higham. 2021. “Reevaluating the Timing of Neanderthal Disappearance in Northwest Europe.” PNAS 118(12): e2022466118. Accessed April 4, 2022. https://doi.org/10.1073/pnas.2022466118.
Dirks, Paul H. G.M., Eric M. Roberts, Hannah Hilbert-Wolf, Jan D. Kramers, John Hawks, and Anthony Dosseto. 2017. “The Age of Homo naledi and Associated Sediments in the Rising Start Cave, South Africa. eLife 6. Accessed April 1, 2022. http://dx.doi.org.offcampus.lib.washington.edu/10.7554/eLife.24231.
Dumasm Laura J., Majesta S O’Bleness, Jonathan M. Davis, C. Michael Dickens, Nathan Anderson, J. G., Keeney, Jay Jackson, Megan Sikela, Armin Raznahan, Jay Giedd, Judith Rapoport, Sandesh S. C. Nagamani, Ayelet Erez, Nicola Brunetti-Pierri, Rachel Sugalski, James R. Lupski, Tasha Fingerlin, Sau Wai Cheung, and James M. Sikela. “DUG1220-Domain Copy Number Implicated in Human Brain-Size Pathology and Evolution.” The American Journal of Human Genetics 91(2): P444-P454. Accessed March 28, 2022. https://www.cell.com/ajhg/fulltext/S0002-9297(12)00373-4.
Girskis, Kelly M., Andrew B. Stergachis, Ellen M. DeGennaro, Ryan N. Doan, Xuyu Qian, Matthew B. Johnson, Peter P. Wang, Gabrielle M. Sejourne, M. Aurel Nagy, Elizabeth A. Pollina, André M. M. Sousa, Taehwan Shin, Connor J. Kenny, Julia L. Scotellaro, Brian M. Debo, Dilenny M. Gonzalez, Lariza M. Rento, Rebecca C. Yeh, Janet H. T. Song, Marc Beaudin, Jean Fan, Peter V. Kharchenko, Nenad Sestan, Michael E. Greenberg, and Christopher A. Walsh. 2021. “Rewiring of Human Neurodevelopmental Gene Regulatory Programs by Human Accelerated Regions.” Neuron 109(20): P3239-P3251. Accessed March 28, 2022. https://doi.org/10.1016/j.neuron.2021.08.005.
Goldfield, Anna. 2019. “The Neanderthal Brain—Clues About Cognintion.” Sapiens. Accessed April 4, 2022. https://www.sapiens.org/column/field-trips/neanderthal-brain/.
Guevara, Elaine E., William D. Hopkins, Patrick R. Hof, John J. Ely, Brenda J. Bradley, and Chet C. Sherwood. 2021. “Comparative Analysis Reveals Distinctive Epigenetic Features of the Human Cerebellum.” PLOS Genetics May 6. Accessed March 28, 2022. https://doi.org/10.1371/journal.pgen.1009506.
Harmand, Sonia, Jason E. Lewis, Craig S. Feibel, Christopher J. Lepre, Sandrine Prat, Arnaud Lenoble, Xavier Boës, Rhonda L. Quinn, Michel Brenet, Adrian Arroyo, Nicholas Taylor, Sophie Clément, Guillaume Daver, Jean-Philip Brugal, Louise Leakey, Richard A. Mortlock, James D. Wright, Sammy Lokorodi, Christopher Kirwa, Dennis V. Kent, and Hélène Roche. 2015. “3.3-Million-Year-Old Stone Tools from Lomekwi 3, West Turkana, Kenya.” Nature 521(7552). Accessed April 1, 2022. http://dx.doi.org.offcampus.lib.washington.edu/10.1038/nature14464.
Hirst, K. Kris. 2019. “Acheulean Handaxe: Definition and History.” ThoughtCo. Accessed April 1, 2022. https://www.thoughtco.com/acheulean-handaxe-first-tool-171238.
Hirst, K. Kris. 2019. “Guide to the Châtelperronian.” ThoughtCo. Accessed April 5, 2022. https://www.thoughtco.com/guide-to-the-chatelperronian-173067.
Hirst, K. Kris. 2019. “Mousterian: A Middle Stone Age Technology That May Be Outmoded.” ThoughtCo. Accessed April 4, 2022. https://www.thoughtco.com/mousterian-definition-167233.
Howard Hughes Medical Institute. 2005. “Evidence That Human Brain Evolution Was a Special Event.” ScienceDaily. Accessed March 28, 2022. https://www.sciencedaily.com/releases/2005/01/050111165229.htm.
Larsen, Clark Spencer. 2014. Our Origins: Discovering Physical Anthropology. New York, NY: W. W. Norton & Company, Inc.
Larsen, Clark Spencer. 2022. Essentials of Biological Anthropology, 5th edition. New York, NY: W. W. Norton & Company, Inc.
Mendez, Fernando L., G. David Poznik, Sergi Castellano, and Carlos D. Bustamante. 2016. “The Divergence of Neandertal and Modern Human Y Chromosomes.” American Journal of Human Genetics 98(4): 728-734. Accessed April 4, 2022. https://doi-org.offcampus.lib.washington.edu/10.1016/j.ajhg.2016.02.023.
Meyer, Matthias, Martin Kircher, Marie-Theres Gansauge, Heng Li, Fernando Racimo, Swapan Mallick, Joshus G. Schraiber, Flora Jay, Kay Prüfer, Cesare de Filippo, Peter H. Sudmant, Can Alkan, Qiaomei Fu, Ron Do, Nadin Rohland, Arti Tandon, Michael Siebauer, Richard E. Green, Katarzyna Bryc, Adrian W. Briggs, Udo Stenzel, Jesse Dabney, Jay Shendure, Jacob Kitzman, Michael F. Hammer, Michael V. Shunkov, Anatoli P. Derevianko, Nick Patterson, Aida M. Andrés, Even E. Eichler, Montgomery Slatkin, David Reich, Janet Kelson, and Svante Pääbo. 2012. “A High-Coverage Genome Sequence from an Archaic Denisovan Individual.” Science 338(6104): 222-226. Accessed April 4, 2022. https://alliance-primo.hosted.exlibrisgroup.com/permalink/f/lvbsh/TN_cdi_pubmedcentral_primary_oai_pubmedcentral_nih_gov_3617501.
Mounier, Aurélien and Marta Mirazón Lahr. 2017. Deciphering African Late Middle Pleistocene Hominin Diversity and the Origin of Our Species. Nature Communications. Accessed November 17, 2021. https://rdcu.be/cBwrm.
Pääbo, Svante. 2015. “The Diverse Origins of the Human Gene Pool.” Nature Reviews Genetics 16: 313-314. Accessed August 21, 2015. http://www.nature.com.offcampus.lib.washington.edu/nrg/journal/v16/n6/full/nrg3954.html.
Park, Michael Alan. 2014. Introducing Anthropology: An Integrated Approach, 6th edition. New York, NY: McGraw-Hill Education.
Paskey Amanda Wolcott and AnnMarie Beasley Cisneros. 2019. “Archaic Homo.” In Explorations: An Open Invitation to Biological Anthropology, edited by Beth Shook, Katie Nelson, Kelsie Aguilera, and Lara Braff. Arlington, VA: American Anthropological Association. Accessed April 1, 2022. https://pressbooks-dev.oer.hawaii.edu/explorationsbioanth/chapter/__unknown__-16/.
Peresane, Marco, Ivana Fiore, Monica Gala, and Antonio Tagliacozzo. 2011. PNAS 108(10): 3888-3893. Accessed April 4, 2022. https://doi.org/10.1073/pnas.1016212108.
Portal, Jane. 2000. Korea: Art and Archaeology. New York, NY: Thames and Hudson.
Raff, Jennifer. 2022. Origin: A Genetic History of the Americas. New York, NY: Twelve.
Reich, David, Richard E. Green, Martin Kircher, Johannes Krause, Nick Patterson, Eric Y. Durand, Bence Viola, Adrian W. Briggs, Udo Stenzel, Philip L. F. Johnson, Tomislav Maricic, Jeffrey M. Good, Tomas Marques-Bonet, Can Alkan, Qiaomei Fu, Swapan Mallick, Heng Li, Matthias Meyer, Evan E. Eichler, Mark Stoneking, Michael Richards, Sahra Talamo, Michael V. Shunkov, Anatoli P. Derevianko, Jean-Jacques Hublin, Janet Kelson, Montgomery Slatkin, and Svante Pääbo. 2010. “Genetic History of an Archaic Hominin Group from Denisova Cave, Sibera.” Nature 468: 1053-1060. Accessed August 21, 2015. http://www.nature.com/nature/journal/v468/n7327/full/nature09710.html.
Richter, Daniel, Rainer Grün, Renaud Joannes-Boyau, Teresa E. Stelle, Fethi Amani, Mathieu Rué, Paul Fernandes, Jean-Paul Raynal, Denis Geraads, Abdelouahed Ben-Ncer, Jean-Jacques Hublin, and Shannon P. McPherron. 2017. “The Age of the Hominin fossils from Jebel Irhoud, Morocco, and the Origins of the Middle Stone Age.” Nature 546: 293-296. Accessed November 17, 2021. https://rdcu.be/cBwrt.
Rightmire, Philip. 2013. “Homo erectus and Middle Pleistocene Hominins: Brian Size, Skull Form, and Species Recognition.” Journal of Human Evolution 65(3): 223-252. Accessed April 1, 2022. https://doi-org.offcampus.lib.washington.edu/10.1016/j.jhevol.2013.04.008.
Rodríquez-Vidal, Joaquín, Francesco d’Errico, Francisco Giles Pacheco, Ruth Blasco, Jordi Rosell, Richard P. Jennigs, Alain Queffelec, Geraldine Finlayson, Darren A. Fa, José Maria Gutiérrez, José S. Carrión, Juan José Negro, Stewart Finlayson, Luís M. Cáceres, Marco A. Bernal, Santiago Fernández Jiménez, and Clive Finlayson. 2014. “A Rock Engraving Made by Neanderthals in Gibraltar.” PNAS 111(37): 13301-13306. Accessed April 4, 2022. https://doi.org/10.1073/pnas.1411529111.
Saneda, Tori and Michelle Field. 2015. “Homo.” Biological Anthropology. Accessed April 5, 2022. https://wikieducator.org/Biological_Anthropology/Unit_3:_Human_Evolution/Homo.
Slatkin, Montgomery and Fernando Racimo. 2016. “Ancient DNA and Human History.” PNAS 113(23): 6380-6387. Accessed April 4, 2022. https://doi.org/10.1073/pnas.1524306113.
Slimak, Ludovic, Clément Zanolli, Tom Higham, Marine Frouin, Jean-Luc Schwenninger, Lee J. Arnold, Martina Demuro, Katerina Douka, Norbert Mercier, Gilles Guérin, Hélène Valladas, Pascale Yvorra, Yves Giraud, Andaine Seguin-Orlando, Ludovic Orlando, Jason E. Lewis, Xavier Muth, Hubert Camus, Ségolène Vandevelde, Mike Buckley, Carolina Mallol, Chris Stringer, and Laure Metz. 2022. “Modern Human Incursion into Neanderthal Territories 54,000 Years Ago at Mandrin, France.” Science Advances 8(6). https://doi-org.offcampus.lib.washington.edu/10.1126/sciadv.abj9496.
Slon, Viviane, Bence Viola, Gabriel Renaud, Marie-Theres Gansauge, Stefano Benazzi, Susanna Sawyer, Jean-Jacques Hublin, Michael V. Shunkov, Anatoly P. Derevianko, Janet Kelson, Kay Prüfer, Matthias Meyer, and Svante Pääbo. 2017. “A Fourth Denisovan Individual.” Science Advances 3(7). Accessed April 4, 2022. https://doi-org.offcampus.lib.washington.edu/10.1126/sciadv.1700186.
Szymik, Brett. 2011. “What’s in Your Brain?” ASU-Ask a Biologist. Accessed March 28, 2022. https://askabiologist.asu.edu/parts-of-the-brain.
Tattersall, Ian and Jeffrey H. Schwartz. 2009. “Evolution of the Genus Homo.” Annual Review of Earth and Planetary Sciences 37: 47-92. Accessed March 30, 2022. https://doi.org/10.1146/annurev.earth.031208.100202.
Thieme, Hartmut. 1997. “Lower Paleolithic Hunting Spears from Germany.” Nature 385(6619): 807-810. Accessed April 4, 2022. https://alliance-primo.hosted.exlibrisgroup.com/permalink/f/lvbsh/TN_cdi_proquest_miscellaneous_78835888
University of Colorado Denver. 2012. “Evolutionary Increase in Size of the Human Brain Explained: Part of a Protein Linked to Rapid Change in Cognitive Ability.” ScienceDaily. Accessed March 28, 2022. https://www.sciencedaily.com/releases/2012/08/120816141537.htm.
Welker, Barbara Helm. 2017. The History of Our Tribe: Hominini. Geneseo, NY: Milne Library Publishing at SUNY Geneseo. Accessed April 4, 2022. https://milneopentextbooks.org/the-history-of-our-tribe-hominini/.
Whitaker, Roger M. Gualtiero B. Colombo, Stuart M. Allen, and Robin I. M. Dunbar. 2016. “A Dominant Social Comparison Hueristic United Alternative Mechanisms for the Evolution of Indirect Reciprocity.” Scientific Reports 6: 31459. Accessed March 28, 2022. https://doi.org/10.1038/srep31459.
Yoshida-Levine, Bonnie. 2019. In Explorations: An Open Invitation to Biological Anthropology, edited by Beth Shook, Katie Nelson, Kelsie Aguilera, and Lara Braff. Arlington, VA: American Anthropological Association. Accessed March 30, 2022. https://pressbooks-dev.oer.hawaii.edu/explorationsbioanth/chapter/__unknown__-15/.
Zhu, Zhao-Yu, Robin Dennell, Wei-Wen Huang, Yi Wu, Zhi-Guo Rao, Shi-Fan Qiu, Jiu-Bing, Xie, Wu Liu, Shu-Qing Fu, Jiang-Wei Han, Hou-Yun Zhou, Ting-Ping Ou Yang, and Hua-Mei Li. 2015. “New Dating of the Homo erectus Cranium from Lantian (Gongwangling), China.” Journal of Human Evolution 78: 144-157. Accessed March 30, 2022. https://doi.org/10.1016/j.jhevol.2014.10.001.

{kind=link}
{kind=link}
{kind=link}
.JPG){kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}